
国家地理标志保护为黎川科技发展,黎川资源开发,黎川面貌改变,黎川绿水青山环境造就,发挥了积极作用。......
中国人爱吃的水稻哪里来?——稻谷多样性之面面观
来源:互联网
2024-11-12 23:24:57 版权归原作者所有,如有侵权,请联系我们

每年的5月22日,联合国环境署将它制定为『国际生物多样性日』,以呼吁保护及永续利用生物多样性的重要性。而2022年的主题『共建地球生命共同体』(Building a shared future for all life) 更是强调:生物多样性是一张巨大的网,将地球上的每个物种紧密的串连起来。那么,稻米的多样性与我们人类是如何串连的?人类的遗传多样性是否也因多元的米食文化而变得丰富呢?
约自1950 年开始,植物学家们大规模且系统性的在世界各地采集稻种,并建立种原库保存种子供科学研究及育种应用。1960年代引领水稻『绿色革命』的奇迹米 IR8,即为张德慈博士与国际稻米研究所 (IRRI) 的育种专家们 (图1),将采集自台湾的半矮型地方栽培种『低脚乌尖』与来自印尼的高茎杆地方种『Peta』杂交,再经过多年繁殖、选拔、评估而育成的高产品种 (图2)。根据国际农业研究谘商组织 (CGIAR) 估计,目前超过78万多种栽培稻与野生稻低温保存在全球40 多个国家的种原库里(图3),不得不令人好奇栽培稻是如何一步一步从野生稻演化出如此丰富的遗传变异及多样性? 考古学家与遗传学家分别从不同的角度收集证据,试图解开这个谜团。

图1. 国际稻米研究所种原库一景。图中人物为张德慈博士,1962 年至1991 年担任国际稻米研究所遗传资源中心主任。

图2. 国际稻米研究所示范田的绿色革命奇迹米 『IR8』 及两亲本『低脚乌尖』与『Peta』出处:IRRI Photo Flickr

图3. 在国际稻米研究所温室内展示的野生稻出处: IRRI Photo Flickr驯化野生稻
时间拉回到一万多年前的欧亚大陆,部分智人 (Homo sapiens) 逐渐放弃狩猎采集的游牧行为,选择耕种畜牧的定居生活,而后发展出『农业』聚落,开始『新石器时代』。那个时期,到处可见带着长芒稻穗(图4)及ㄧ碰就落粒的各种野生稻族群。有些野生稻喜欢生长在潮湿多雨的阴凉河谷区,有些则比较适应干燥的土壤及温暖的气候。部分野生稻发展出多年生的生长习性,依赖地下匍匐茎繁殖,种子则进入休眠,等环境条件对了才发芽。有些则是单年生,可以在风或虫的帮忙下完成异花授粉(图5)。有趣的是,许多野生稻的表征或特性,在现代广泛种植的亚洲栽培稻几乎都看不见了。但是,控制这些特性的遗传因子仍有可能隐藏在古老的地方种 (landraces) 或野生近缘种的基因池 (gene pool) 内,等待我们去发掘!

图4. 具长芒的野生稻种子出处:IRRI Photo Flickr

图5. 野生稻雄蕊花药明显外露,易促成异花授粉。出处:IRRI Photo Flickr
考古学家、人类学家与遗传学家非常好奇亚洲栽培稻 (Oryza sativa) 是在什么时候出现,在哪里出现?一万多年前的野生稻,它是如何被人类『驯化』成适合耕种、容易收获保存的稻种?考古学家四处挖掘遗址,藉由分析碳同位素或陶器残骸内的稻硅酸体 (phytolith)推估年代。从部分碳化米的外观型态,可约略区分野生型或栽培型,但大多数出土的稻谷遗骸因年代久远而不易鉴别粒型。科学家进一步利用高倍率显微镜观察遗址出土的稻谷底部与穗柄轴之间的离层带 (abscission zone) 构造,野生稻因为形成离层带而容易落粒(图6E, 6H),驯化稻却无明显的离层带(图6D, 6G),此特征可被用来判定水稻驯化的程度。相较于不易管理的常落粒野生稻,驯化稻谷成熟后仍与穗柄轴相连,早期的农夫可用石器工具在适当的时间统一采收,大大的提高农作效率及收获产量,也因此加速驯化稻的普及性。目前许多亚洲国家包括台湾的考古遗址都有驯化稻的踪迹,然而只有少数如中国的河姆渡/田螺山遗址仍可发现不同比例的野生稻与驯化稻遗骸,推估6600-6900年前在长江中下游流域的水稻已完成驯化。位于长江下游的上山遗址,透过当地出土的稻谷遗骸硅酸体定年结果,更将栽培稻的驯化初期追溯至9400年前,是目前发现最早的驯化稻。那么,长江流域是唯一的驯化稻起源地吗?
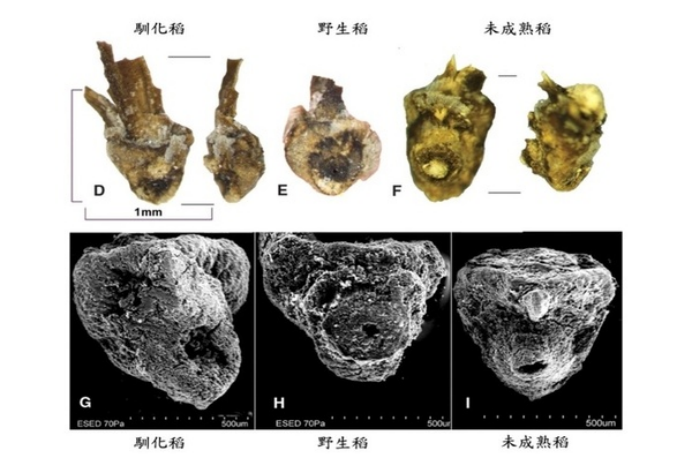
图6. 田螺山遗址出土的野生稻与驯化稻谷基部构造。野生稻因为形成离层带而容易落粒(图6E, 6H),驯化稻则无明显的离层带 (图6D, 6G)。改编自出处:Fig. 3 in Fuller et al (2009) Science 323, 1607-1610
栽培稻是单一起源还是多重起源?
相较于非洲栽培稻 (Oryza glaberrima),亚洲栽培稻 (Oryza sativa) 是当今全世界栽种面积最广的稻种,可依据型态、分布地理位置、遗传背景等特性,分出黏度高、谷粒短圆、生长于温带及亚热带的粳稻 (japonica rice) 与黏度低、谷粒瘦长、多种植于热带及亚热带的籼稻 (indica rice)。有趣的是,当遗传学家将粳稻与籼稻杂交后,发现有些后代会产生不稔实 (sterile) 的种子,因此推测在驯化及之后的传播过程中,粳稻与籼稻的基因序列变得越来越不一样而渐渐形成生殖隔离,故将其视为两个亚种 (subspecies)。那么,粳稻与籼稻是如何从一万多年前的野生稻驯化而来的呢?哪一个先出现?或是两者同时在不同的地方被驯化的?亦或还有其他的驯化路径?考古研究虽能推测水稻驯化的时间及地点,但是较难从数千年前出土的稻谷遗骸型态判断驯化稻是粳稻或籼稻,也不易从碳化稻萃取出完整的DNA 分子进行分析比较。于是,遗传学家从各地采集存活至今的野生稻、野生近缘种及地方栽培稻,试图从它们的DNA序列变化,探索水稻早期的驯化足迹及传播路径。前段提到,失去落粒性是一个非常重要的驯化性状,当科学家比较野生稻及栽培稻控制落粒程度的主要基因 (sh4) 序列,发现所分析的栽培稻,不论是籼稻或粳稻,都具有相同的遗传讯息,跟野生稻完全不同,因此推论水稻驯化是单一起源。除了落粒性,还有一些驯化性状如株型、分櫱程度的基因分析也得到类似的结果。然而,部分科学家认为以少数几个驯化基因的序列变化来支持水稻驯化是单一起源的证据太薄弱,无法被完全说服。近年来,科学家藉由分析大量野生稻及栽培稻的全基因体序列,推测一万多年前的人类在广西附近的珠江流域,开始将当地的野生稻族群驯化出粳稻。粳稻随后往外漂移,有的往中国北方及韩国、日本移动,有的往南方传播,进入东南亚地区,与当地原有的野生稻种杂交,开始第二次驯化,产生籼稻。虽然最新的DNA分子演化证据与考古稻谷遗骸证据对驯化稻起源地的推论有所出入,科学家们仍努力不懈的继续应用新技术及新方法探索这个有趣的问题。水稻多样性
水稻驯化是一个漫长且复杂的过程,在这几百年甚至几千年的时间里,从初期的野生族群与驯化中的族群共同存在,到后来半驯化或完全驯化的稻种随着人类的迁徙传播至各地,适应各种多元的地理环境,发展出今日丰富的多样性(图7)。现今全世界栽培稻的耕种面积约有1亿6千多公顷,90% 以上分布在亚洲。我们可以在喜马拉雅山脚下的不丹看到水稻梯田,也可以在湿热的印度及孟加拉的恒河三角洲发现深水稻,北至苏联、日本北海道,南至中南美洲、澳洲,都可以种稻。栽培稻为了适应不同环境条件,有的对日照长度从敏感变钝感;有的根系特别发达可于旱地生长;有的长期种植在高纬度或高海拔地区,因此特别耐低温;有的则发展出耐热生理机制,可抵抗高温的冲击。自从绿色革命以来,高产的商业品种或推广品种深受多数农民的青睐,因此,传统的地方种渐渐从农田消失,生物多样性也随之下降。幸运的是,至今还有些农夫仍种着他们的祖先世世代代传下来的稻种,这些被称为传家宝 (heirloom) 的地方栽培种,虽然耕种面积不大,产量也不出色,但是却能丰富水稻的遗传多样性,也对当地的历史、文化、风土民情的保存扮演重要的角色,因此,保种是一场迫在眉睫的全球行动。

图7. 自世界各地搜集的地方栽培稻种原出处:IRRI Photo Flickr
国际稻米研究所的科学家们调查3000 多个来自世界各地的栽培稻型态如株高及粒型、生长习性如开花时间、分櫱数以及稻米质量等多个性状,丰富的多样性也反映在稻种的基因序列里面。科学家比较这3000 多个栽培稻的全基因组信息,发现超过三千多万个DNA分子变化,包含单一硷基差异、小片段的插入或缺失,大片段的结构变化等等。演化亲缘关系分析除了鉴别出籼稻、粳稻两大分支外,籼稻族群内可依地理位置分出东亚籼稻、南亚籼稻、东南亚籼稻及一小群来自于孟加拉的夏稻 (aus) (图8);粳稻族群内则可再区分出温带型 (temperate)、热带型 (tropical) 、亚热带型 (subtropical) 及香米 (aromatic) 等数个次族群,温带型粳稻多分布于东亚如日本、韩国,中国北方等地、热带及亚热带粳稻则分布于东南亚地区。大家所熟悉的爪哇型稻 (Javanica rice) 则多属于热带或亚热带粳稻。台湾地理位置特殊,育种家因应消费需求育成各种类型的米种,配合农民丰富的栽培管理经验,不论是栽种面积最广的粳稻如台南11号、媲美越光米的台粳9号、台南16号,或是香米如高雄147号、台中194号,以及籼米如台中籼10号,均能在这片土地上开花结果。
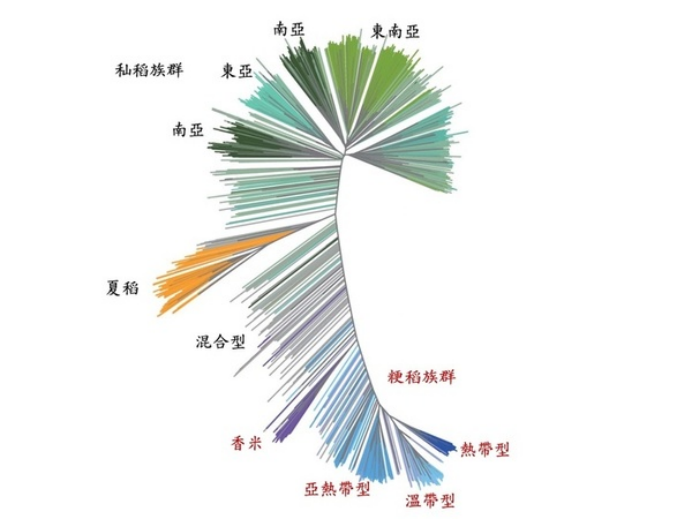
图8. 3010个栽培稻的亲缘关系树改编自出处:Fig.1 in Wang, W., Mauleon, R., Hu, Z. et al. (2018) Nature 557, 43–49
透过多年来的科学研究,我们可以清楚看见生态环境及人为选拔影响稻种遗传多样性的丰富度。近年来,育种家们大规模的探勘及评估水稻种原的遗传歧异度及各种特性,孟加拉的夏稻就被发现特别适合在高温多雨的季节生长,而且对多种逆境的抵抗能力也很好。育种学家已经开始利用夏稻的优良特性改良品种,最有名的例子即为国际稻米研究所从非常耐淹水的夏稻品种FR13A找到抗性基因 Sub1,再利用传统杂交配合分子标志辅助选育方法 (非基因改造),将FR13A的耐淹基因Sub1导入原本不耐淹的地方适应品种使其获得抗性(图9),目前已在东南亚国家大规模的推广种植。
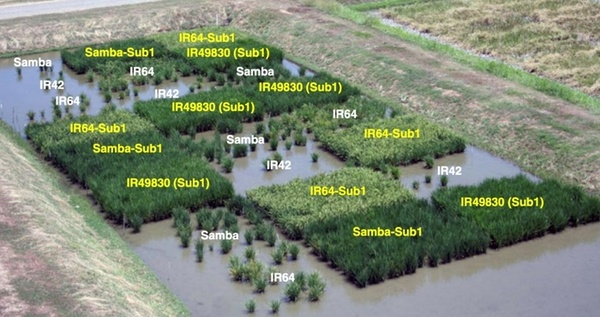
图9. 国际稻米研究所的田间试验,显示导入耐淹基因Sub1 的品种如Samba-Sub1在淹水 (1.25 公尺深) 17天后的表现远优于原本没有耐淹基因的亲本 Samba。出处:Bailey-Serres, J., Fukao, T., Ronald, P. et al. (2010) Rice 3, 138–147
稻米的营养成分以淀粉为主,粳米与籼米在烹煮后,低直链淀粉的粳米黏度高,反之,高直链淀粉的籼米较松散。籼粳直链淀粉含量的差异,影响米质及口感,因此,人类在不同的地理环境,发展出多元的米食文化。为了能有效率的将淀粉代谢成能量,人类在演化过程中,产生数目不等的淀粉酶基因 (amylase, AMY) 消化淀粉,遗传学家发现人类族群间存在着淀粉酶基因数目的差异,而且基因数目的多寡与人类食用淀粉的偏好性有高度的相关性,但是否与粳米或籼米的选择有关仍有待探索。
结语
经过时间洗礼而存活下来的每个物种就象是一颗颗时空胶囊,它们的DNA 里记录着过去所发生的突变、天择、人择、移地适应及遗传物质重组、互换、交流等等事件,这些事件也正是形塑生物多样性的原动力。因此,不论是低温保存在种原库里的稻种,或是正随着不同季节在各种生态、地理环境下开花结穗的稻子,都需要我们好好的保护及珍惜它们,不只是为了我们人类的生存,也为了地球这一张巨大的网所串起的所有生物。
参考文献
[1] 永远的DGWG https://epaper.ntu.edu.tw/view.php?id=21775
[2] 寻回水稻生物多样性 https://sa.ylib.com/MagArticle.aspx?id=5183
[3] The Domestication Process and Domestication Rate in Rice: Spikelet Bases from the Lower Yangtze (2009) https://www.science.org/doi/full/10.1126/science.1166605
[4] Rice Domestication by Reducing Shattering (2006) https://www.science.org/doi/10.1126/science.1123604
[5] A map of rice genome variation reveals the origin of cultivated rice (2012) https://www.nature.com/articles/nature11532
[6] Genomic variation in 3,010 diverse accessions of Asian cultivated rice (2018) https://www.nature.com/articles/s41586-018-0063-9
[7] Diet and the evolution of human amylase gene copy number variation (2007)https://www.nature.com/articles/ng2123
以上内容由办公区教程网摘抄自中国科普网可供大家参考!
上一篇:43年 就为了一头牛? 下一篇:开远市农函大开展水稻高产种植管理培训
相关文章

